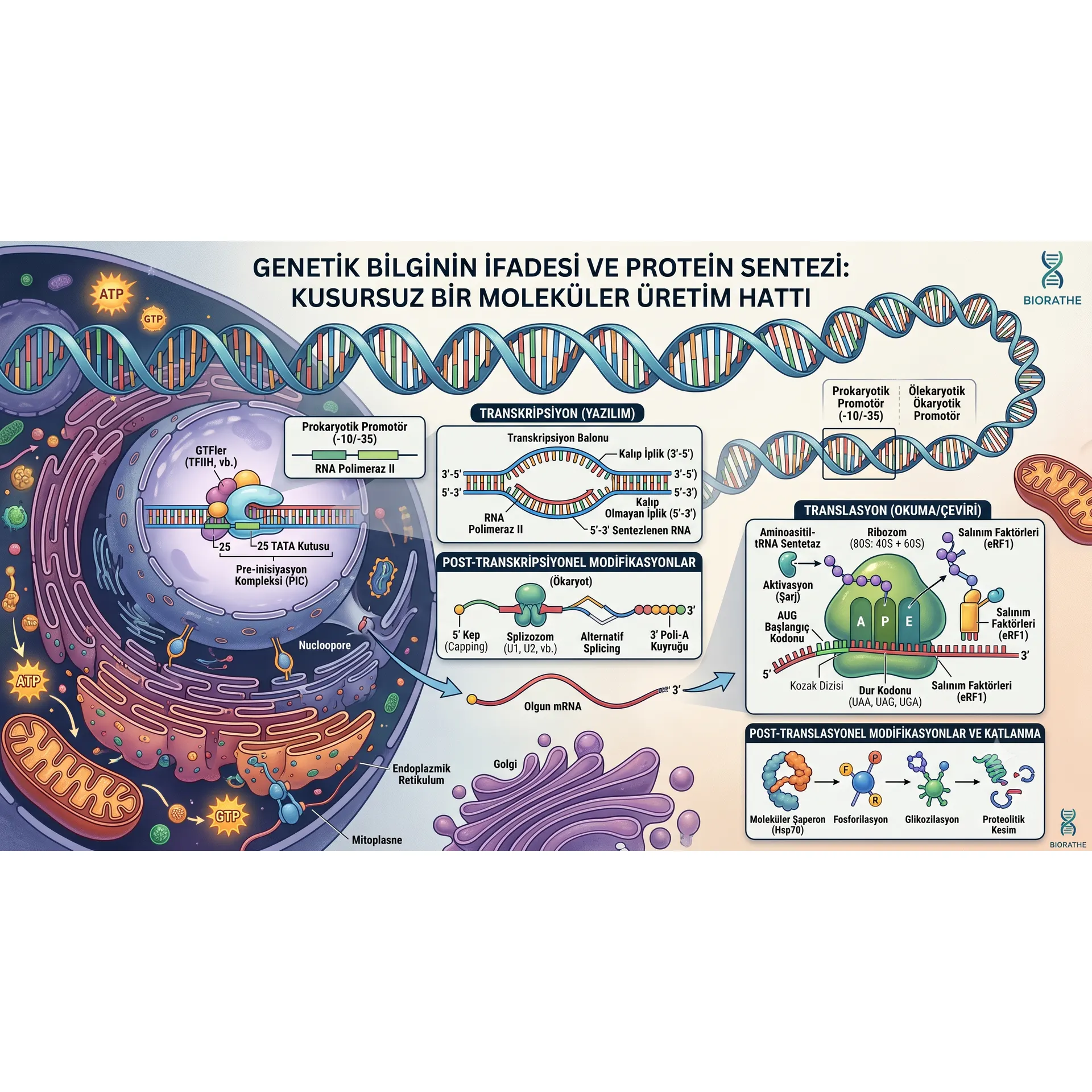
Moleküler biyolojinin ve genetiğin temel dogması (Central Dogma), genetik bilginin yönünü çift iplikli DNA molekülünden fonksiyonel protein yapılarına doğru çizer. Hücre içindeki tüm metabolik, yapısal ve sinyal iletim süreçlerinin merkezinde yer alan proteinlerin sentezi, rastgele bir birleşme değil; hata payı neredeyse sıfıra indirilmiş, kusursuz regüle edilen moleküler bir üretim hattıdır. Bu üretim hattı temel olarak iki büyük ve karmaşık aşamadan oluşur: Transkripsiyon (Yazılım) ve Translasyon (Okuma/Çeviri).
Bu yazıda, ökaryotik ve prokaryotik organizmalar arasındaki nüanslara da değinerek, protein sentezinin mekanik adımlarını, görev alan enzim sistemlerini ve post-transkripsiyonel/post-translasyonel modifikasyonları lisans düzeyinde bir derinlikle ele alacağız.
Transkripsiyon: Genetik Kodun Kalıptan Çıkarılması
Transkripsiyon, DNA üzerinde lokalize olan spesifik bir gen dizisinin, RNA Polimeraz enzimi ve yardımcı transkripsiyon faktörleri aracılığıyla tamamlayıcı (komplementer) bir tek iplikli RNA molekülüne (primer transkript) dönüştürülmesi sürecidir. Süreç üç ana evrede incelenir: Başlama, Uzama ve Sonlanma.
Başlama (İnisiyasyon)
Transkripsiyonun başlaması, genomun hangi bölgesinin, ne zaman ve ne kadar oranla ifade edileceğinin belirlendiği ana kontrol noktasıdır. Süreç, RNA Polimeraz enziminin DNA üzerindeki promotör adı verilen özel düzenleyici dizilere bağlanmasıyla başlar.
- Prokaryotlarda: Tek bir RNA Polimeraz türü bulunur. Enzimin promotörü tanıyabilmesi için çekirdek enzime (core enzyme) Sigma faktörü adı verilen geçici bir alt birimin bağlanması gerekir. Bu birleşme ile oluşan yapıya holoenzim denir. Prokaryotik promotörlerde yer alan -10 (Pribnow Kutusu / TATAAT) ve -35 (TTGACA) konsensüs dizileri, sigma faktörü tarafından tanınır.
- Ökaryotlarda: Süreç çok daha karmaşıktır. Üç farklı RNA Polimeraz bulunur (RNA Pol I: rRNA, RNA Pol II: mRNA ve snRNA, RNA Pol III: tRNA ve 5S rRNA sentezler). Protein kodlayan genlerden sorumlu olan RNA Polimeraz II, promotör bölgeye (genellikle TATA kutusu, -25 lokalizasyonunda) doğrudan bağlanamaz. Öncelikle Genel Transkripsiyon Faktörleri (GTF'ler: TFIID, TFIIB, TFIIF, TFIIE, TFIIH) promotör bölgeye sırayla yerleşerek bir Pre-inisiyasyon Kompleksi (PIC) oluşturur. TFIIH'ın kinaz aktivitesi ile RNA Pol II'nin karboksil terminal ucu (CTD) fosforillenir, bu da enzimin promotörden salınarak uzama fazına geçmesini sağlar
RNA Polimeraz, promotör dizisine kararlı bir şekilde bağlandıktan sonra, DNA çift sarmalının yaklaşık 12-14 baz çiftlik lokal bir bölgesini hidrole ederek açar. Bu açık yapıya transkripsiyon balonu adı verilir.
Uzama (Elongasyon)
Transkripsiyon balonu oluştuktan sonra, RNA Polimeraz DNA'nın kalıp ipliğini (template strand, 3' - 5' yönünde) okumaya başlar. Yeni RNA zinciri ise her zaman 5 - 3' yönünde sentezlenir.
Enzim, ortamda bulunan ribonükleosit trifosfatları (ATP, UTP, GTP, CTP) kalıp iplikteki pürin ve pirimidin bazlarına komplementer olacak şekilde (A karşısına U, T karşısına A, G karşısına C, C karşısına G) phosphodiester bağlarıyla birbirine bağlar. Sentez sırasında geçici bir RNA-DNA hibrit sarmalı oluşur. RNA Polimeraz ilerledikçe, arkasında kalan DNA sarmalı yeniden kapanır ve sentezlenen RNA ucu transkripsiyon balonundan dışarı doğru sarkar.
Sonlanma (Terminasyon)
RNA Polimerazın gen sonundaki spesifik dizilere ulaşmasıyla transkripsiyon sonlanır ve RNA transkripti serbest kalır.
Prokaryotik Sonlanma: İki mekanizma ile gerçekleşir.
- Rho-bağımsız (İntrinsik) Sonlanma: Yeni sentezlenen RNA üzerinde peş peşe gelen GC zengin bir bölge, kendi içinde katlanarak bir saç tokası (hairpin) yapısı oluşturur. Bu yapının hemen ardından gelen poli-U kuyruğu, DNA'daki adeninler ile zayıf ikili hidrojen bağları kurduğu için transkripsiyon kompleksi dayanıksızlaşır ve disosiye olur.
- Rho-bağımlı Sonlanma: Rho adı verilen bir ATPaz helikaz proteini, RNA üzerindeki rut dizisine bağlanır, RNA Polimerazın arkasından ilerler ve enzim duraklama noktasına geldiğinde RNA-DNA hibrit yapısını mekanik olarak çözer.
Ökaryotik Sonlanma
RNA Pol II, poliadenilasyon sinyal dizisini (AAUAAA) sentezledikten sonra, transkript özel endonükleazlar tarafından kesilir. Kalan RNA ucu exonükleazlar (Xrn2) tarafından sindirilir ve RNA Pol II DNA'dan uzaklaştırılır (Torpedo modeli).
Post-Transkripsiyonel Modifikasyonlar (RNA İşlenmesi)
- Prokaryotlarda transkripsiyon ve translasyon aynı hücresel kompartımanda (sitoplazmada) eş zamanlı olarak gerçekleşir; mRNA sentezlenirken bir yandan ribozomlar tarafından okunmaya başlar. Ancak ökaryotlarda transkripsiyon çekirdekte, translasyon sitoplazmada gerçekleştiği için, sentezlenen primer transkriptin (pre-mRNA) sitoplazmaya transfer edilmeden önce korunması ve olgunlaştırılması gerekir.
- 5' Kep (Capping) Modifikasyonu: Transkripsiyonun henüz başlarında (yaklaşık 20-30 nükleotit sentezlendikten sonra), pre-mRNA'nın 5' ucuna atipik bir 5'-5' trifosfat bağı ile 7-metilguanozin molekülü eklenir. Bu yapı, mRNA'yı hücresel 5' - 3' ekzonükleaz yıkımından korur ve translasyon aşamasında ribozomun mRNA'yı tanımasına yardımcı olur.
- Poliadenilasyon (3' Poli-A Kuyruğu): Sentezlenen pre-mRNA'nın 3' ucu kesildikten sonra, Poli-A Polimeraz (PAP) enzimi tarafından şablon kullanmaksızın 150-250 adet adenin nükleotiti eklenir. Poli-A kuyruğu, mRNA'nın kararlılığını (stabilitesini) sağlar ve çekirdekten sitoplazmaya çıkış vizesidir.
- RNA Splicing (Ekleme): Ökaryotik genler kesintili yapılardır; protein kodlayan ekzon dizileri ile protein kodlamayan intron dizilerini bir arada içerirler. Splizozom (Splicosome) adı verilen büyük bir ribonükleoprotein kompleksi (snRNP'ler: U1, U2, U4, U5, U6), intron dizilerini ilmek oluşturarak (lariat yapısı) kesip çıkarır ve ekzonları hassas bir şekilde birbirine bağlar. Bazı durumlarda alternatif splicing mekanizması devreye girerek, tek bir pre-mRNA dizisinden farklı ekzon kombinasyonları ile birden fazla varyant protein sentezlenebilir.
Translasyon: Nükleik Asit Dilinden Amino Asit Diline Çeviri
Olgunlaşan mRNA, nükleer gözeneklerden sitoplazmaya geçtikten sonra translasyon süreci başlar. Bu aşamada 4 harfli nükleotit dili (A, U, G, C), 20 harfli amino asit diline çevrilir. Sürecin temel bileşenleri mRNA, tRNA, Ribozomlar ve Aminoasitil-tRNA Sentetaz enzimleridir.
Genetik Kodun Özellikleri
mRNA üzerindeki üçlü nükleotit dizilerine kodon denir. Toplam 64 kodon bulunur. Bunlardan AUG (Metiyonin) evrensel başlangıç kodonudur. Üç adet kodon ise (UAA, UAG, UGA) stop kodonudur ve amino asit karşılıkları yoktur. Genetik kod dejeneredir (redandant); yani birden fazla farklı kodon, aynı amino asidi şifreleyebilir (Örn: Lösin için 6 farklı kodon vardır). Ancak kod kesindir (unambiguous); bir kodon asla birden fazla farklı amino asidi şifrelemez.
tRNA'ların Aktivasyonu (Aminoasitilasyon)
tRNA molekülleri, antikodon döngüleri ile mRNA'daki kodonları tanırken, 3' akseptör kollarına doğru amino asidi taşımak zorundadır. Bu eşleşmeyi hatasız yapan enzim ailesi Aminoasitil-tRNA Sentetazlardır. Her amino asit için spesifik olan bu enzimler, ATP enerjisini kullanarak amino asidi tRNA'nın 3'-OH ucuna ester bağı ile bağlar. Bu sürece tRNA'nın "şarj edilmesi" veya aktivasyonu denir.
Ribozomun Anatomisi
Ribozomlar, rRNA ve proteinlerden oluşan devasa ribozim kompleksleridir. Küçük ve büyük olmak üzere iki alt birimden oluşurlar (Prokaryotlarda 30S ve 50S = 70S; Ökaryotlarda 40S ve 60S = 80S). Büyük alt birim üzerinde tRNA'ların konumlandığı üç kritik bölge bulunur:
- A Bölgesi (Aminoasitil sites): Yeni gelecek olan şarjlı tRNA'nın giriş yaptığı kabul alanı.
- P Bölgesi (Peptidil sites): Uzayan polipeptit zincirini taşıyan tRNA'nın konumlandığı merkez.
- E Bölgesi (Exit sites): Amino asidini bırakıp yüksüz kalan tRNA'nın ribozomu terk ettiği çıkış kapısı.
Translasyonun Adımları
1. Başlama (İnisiyasyon)
- Prokaryotlarda: Küçük alt birim (30S), mRNA üzerindeki başlangıç kodonunun hemen yukarısında yer alan Shine-Dalgarno (AGGAGG) dizisine bağlanarak doğru okuma çerçevesini (reading frame) bulur. İlk amino asit modifiye edilmiş bir metiyonindir: Formil-metiyonin (fMet-tRNAfMet). İnisiyasyon faktörleri (IF1, IF2, IF3) eşliğinde büyük alt birim de komplekse katılır.
- Ökaryotlarda: Küçük alt birim (40S), eIF4F kompleksi ile mRNA'nın 5' kep yapısını tanır ve mRNA boyunca 5' - 3' yönünde tarama (scanning) yaparak uygun konsensüs dizisi içindeki (Kozak Dizisi) AUG başlangıç kodonunu bulur. İlk tRNA doğrudan Met-tRNAiMet olarak P bölgesine yerleşir. Ardından 60S büyük alt birimi bağlanarak fonksiyonel 80S kompleksi kurulur.
2. Uzama (Elongasyon)
P bölgesinde başlangıç tRNA'sı bulunurken, bir sonraki kodona uygun olan yeni bir aminoasitil-tRNA, uzama faktörleri (Prokaryotlarda EF-Tu, Ökaryotlarda eEF1A) ve GTP hidrolizi yardımıyla A bölgesine giriş yapar.
Büyük alt birimin katalitik merkezinde yer alan Peptidil Transferaz aktivitesi (bu aktiviteyi doğrudan 23S/28S rRNA yürütür, yani bir ribozimdir), P bölgesindeki tRNA'nın ucunda bulunan amino asit ile A bölgesindeki tRNA'nın taşıdığı amino asit arasında bir peptit bağı kurar. Bu esnada polipeptit zinciri tamamen A bölgesindeki tRNA'nın üzerine transfer edilmiş olur.
Hemen ardından, translokasyon faktörleri (Prokaryotlarda EF-G, Ökaryotlarda eEF2) eşliğinde ribozom, mRNA üzerinde tam olarak 3 nükleotit (bir kodon) ileri kayar. Bu hareketle:
- P bölgesindeki boşalmış tRNA E bölgesine geçer ve salınır.
- A bölgesindeki peptit zincirini taşıyan tRNA P bölgesine kayar.
- A bölgesi yeni bir tRNA'yı kabul etmek üzere tekrar boşalır.
3. Sonlanma (Terminasyon)
Ribozomun A bölgesine üç durdurma kodonundan biri (UAA, UAG, UGA) geldiğinde süreç durur. Hücrede bu kodonları tanıyacak bir tRNA molekülü yoktur. Bunun yerine A bölgesine Salınım Faktörleri (Release Factors: RF1/RF2 veya ökaryotik eRF1) bağlanır.
alınım faktörleri, peptidil transferaz merkezini uyararak polipeptit zinciri ile son tRNA arasındaki ester bağının su molekülü kullanılarak hidroliz edilmesini sağlar. Tamamlanan protein zinciri sitoplazmaya salınır; ribozom alt birimleri, mRNA ve salınım faktörleri bir sonraki sentez döngüsünde kullanılmak üzere tamamen disosiye olur.
4. Post-Translasyonel Modifikasyonlar ve Katlanma
Ribozomdan taze çıkmış bir polipeptit zinciri, henüz fonksiyonel bir protein değildir (primer yapıdadır). Proteinin üç boyutlu aktif konformasyonunu (sekonder, tersiyer, kuaterner yapılar) kazanabilmesi için düzgün katlanması ve kimyasal olarak modifiye edilmesi gerekir.
- Moleküler Şaperonlar: Hücre içinde yeni sentezlenen proteinlerin yanlış katlanmasını veya agregasyon (çökelme) oluşturmasını engelleyen, katlanma sürecine refakat eden Hsp70 ve Şaperonin (GroEL/ES complexes) gibi özel proteinlerdir.
- Kovalent Modifikasyonlar: Proteinin aktivitesini, lokalizasyonunu veya ömrünü belirlemek için yan zincirlere fonksiyonel gruplar eklenir:
Fosforilasyon: Kinaz enzimleri tarafından Serin, Treonin veya Tirozin kalıntılarına fosfat eklenmesi (sinyal iletiminin açılıp kapanması).
Glikozilasyon: Endoplazmik retikulum ve Golgi aygıtında proteine karbonhidrat zincirlerinin eklenmesi (özellikle membran ve salgı proteinleri için kritik).
Proteolitik Kesim: Sentezlenen inaktif öncül proteinlerin (Örn: Proinsülin) belirli bölgelerinden kesilerek aktif forma (İnsülin) dönüştürülmesi.
Referanlar
- Alberts, B., Heald, R., Johnson, A., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular biology of the cell (7th ed.). W. W. Norton & Company.
- Crick, F. (1970). Central dogma of molecular biology. Nature, 227(5258), 561–563. https://doi.org/10.1038/227561a0
- Ibba, M., & Söll, D. (2000). Aminoacyl-tRNA synthesis. Annu. Rev. Biochem., 69(1), 617–650. https://doi.org/10.1146/annurev.biochem.69.1.617
- Kozak, M. (2005). Regulation of translation via mRNA structure in prokaryotes and eukaryotes. Gene, 361, 13–37. https://doi.org/10.1016/j.gene.2005.06.014
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Martin, K. C. (2021). Molecular cell biology (9th ed.). W. H. Freeman and Company.
- Plotkin, J. B., & Kudla, G. (2011). Codon usage bias: Mutations, selection and pathogenesis. Nature Reviews Genetics, 12(1), 32–42. https://doi.org/10.1038/nrg2899
- Rodnina, M. V., & Wintermeyer, W. (2001). Fidelity of aminoacyl-tRNA selection on the ribosome. Annu. Rev. Biochem., 70(1), 415–435. https://doi.org/10.1146/annurev.biochem.70.1.415
- Shatkin, A. J. (1976). Capping of eucaryotic mRNAs. Cell, 9(4), 645–653. https://doi.org/10.1016/0092-8674(76)90128-8
- Sharp, P. A. (1994). Split genes and RNA splicing. Cell, 77(6), 805–815. https://doi.org/10.1016/0092-8674(94)90130-9
- Zaher, H. S., & Green, R. (2009). Fidelity at the molecular level: Lessons from protein synthesis. Cell, 136(4), 746–762. https://doi.org/10.1016/j.cell.2009.01.049




Henüz yorum yapılmadı. İlk yorumu sen bırakabilirsin.