Moving Beyond the "Scissors"
The CRISPR-Cas9 revolution provided the scientific community with a programmable "molecular scissor." However, as research progressed into clinical phases, the limitations of inducing Double-Strand Breaks (DSBs) became evident. DSBs often trigger uncontrolled insertions or deletions (indels) via the Non-Homologous End Joining (NHEJ) pathway, which can lead to genotoxicity or unintended gene knockouts. For researchers focused on precision medicine, the goal has shifted from "cutting" to "chemical rewriting." This article explores the biochemical nuances of Base Editing and Prime Editing—two technologies that are redefining the boundaries of genomic stability and therapeutic efficacy.
The Molecular Logic of Base Editing
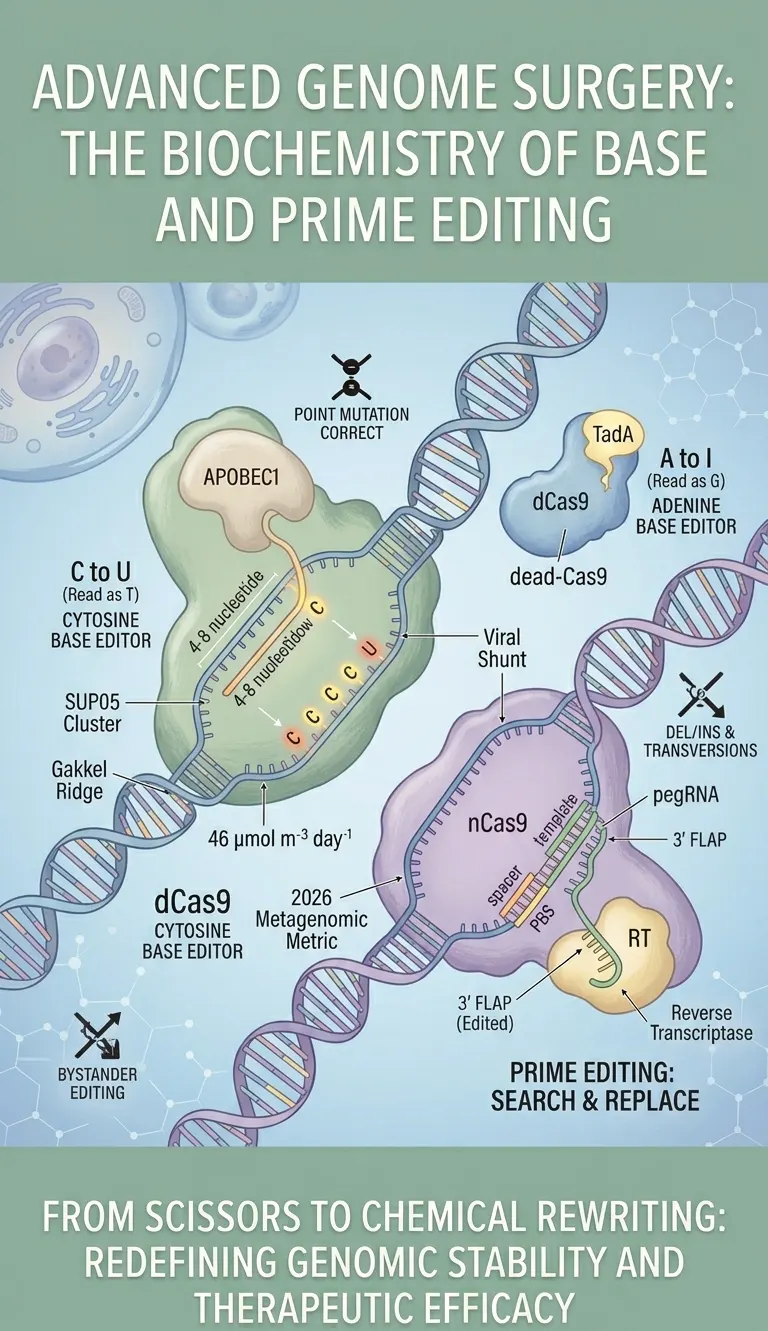
Base editors are chimeric proteins that combine the targeting capability of a "dead" or "nickase" Cas9 (dCas9/nCas9) with the catalytic power of nucleoside deaminases. There are two primary classes: Cytosine Base Editors (CBEs) and Adenine Base Editors (ABEs).
CBEs utilize a cytidine deaminase (such as APOBEC1) to convert Cytosine (C) to Uracil (U). Within the cellular context, Uracil is read as Thymine (T) during DNA replication. To prevent the cell's natural Uracil DNA Glycosylase (UNG) from repairing the U back to C, these editors are often fused with a Uracil Glycosylase Inhibitor (UGI). On the other hand, ABEs use an engineered deoxyadenosine deaminase (TadA) to convert Adenine (A) to Inosine (I), which the cellular machinery interprets as Guanine (G).
The academic importance of base editing lies in its ability to correct "Point Mutations"—which account for roughly 60% of human pathogenic genetic variants—without the risk of chromosomal translocations associated with DSBs. However, "bystander editing" remains a technical hurdle. If multiple Cs or As are present within the "activity window" (typically 4-8 nucleotides), the enzyme may modify all of them, potentially creating new synonymous or non-synonymous mutations.
Prime Editing: The Versatile "Search-and-Replace" System
In 2019, the development of Prime Editing by the Liu Lab introduced a paradigm shift. Unlike base editors, which are restricted to specific transitions (C→T or A→G), Prime Editors can perform all twelve possible base-to-base conversions, as well as targeted insertions and deletions.
The Prime Editing complex consists of an nCas9 fused to an engineered Reverse Transcriptase (RT) and a multi-functional "prime editing guide RNA" (pegRNA). The pegRNA is the "mastermind" of the operation: it contains a spacer sequence to locate the target, a primer binding site (PBS) to anchor the nicked DNA, and a template sequence containing the desired edit.
When the nCas9 nicks the "PAM-strand," the PBS of the pegRNA hybridizes to the target DNA. The RT then extends the DNA strand according to the pegRNA template. This creates a "3' flap" containing the edit and a "5' flap" containing the original sequence. Through a process of flap equilibration and cellular DNA repair, the edited sequence is incorporated into the genome. Because Prime Editing does not require a donor DNA template or DSBs, it is significantly more efficient in primary cells and in vivo models compared to traditional Homology-Directed Repair (HDR).
Navigating the "Off-Target" Landscape
For any academician working in gene therapy, "off-target" effects are the primary concern. Base editors can sometimes induce RNA-level deamination, where the deaminase acts on cellular RNA instead of the target DNA. Prime Editing, due to its requirement for three separate hybridization events (spacer-DNA, PBS-DNA, and flap-DNA), is theoretically more specific than classical CRISPR.
However, the size of these tools is a logistical challenge. A Prime Editor is a massive protein-RNA complex, often exceeding the carrying capacity of standard Adeno-Associated Virus (AAV) vectors. Researchers are currently investigating "split-intein" delivery systems, where the editor is divided into two halves and reconstituted only inside the target cell. Furthermore, the use of "circularized pegRNAs" (cpegRNAs) has recently been shown to increase the stability of the guide RNA, thereby enhancing editing efficiency in difficult-to-reach tissues like the central nervous system.
The Path to Clinical Translation
As we move toward "CRISPR 3.0," the focus is on "PAM-less" or "PAM-flexible" Cas variants (like SpRY). These engineered proteins can recognize almost any sequence in the genome, removing the restriction that the target must be adjacent to a specific NGG motif. This level of precision allows us to target regulatory elements, enhancers, and deep intronic mutations that were previously inaccessible.
For the PhD student or the PI, the choice between Base and Prime editing depends on the specific biological question. Base editing offers higher efficiency for simple transitions, while Prime editing offers unparalleled flexibility for complex indels. The future of the field lies in the hybrid use of these tools, perhaps using Base editors for metabolic silencing and Prime editors for structural gene correction.
References
1. Anzalone, A. V., et al. (2019). Search-and-replace genome editing without double-strand breaks or donor DNA. Nature, 576(7785), 149-157.
2. Gaudelli, N. M., et al. (2017). Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature, 551(7681), 464-471.
3. Komor, A. C., et al. (2016). Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature, 533(7603), 420-424.
4. Rees, H. A., & Liu, D. R. (2018). Base editing: Precision chemistry on the genome and transcriptome of living cells. Nature Reviews Genetics, 19(12), 770-788.
5. Kantor, A., et al. (2020). CRISPR-Cas9, Cas12a, and Cas13a: A guide to mastery. Molecular Therapy, 28(11), 2312-2325.
6. Newby, G. A., & Liu, D. R. (2021). In vivo somatic cell gene editing with base editors and prime editors. Trends in Genetics, 37(12), 1111-1124.


No comments yet. You can leave the first one.