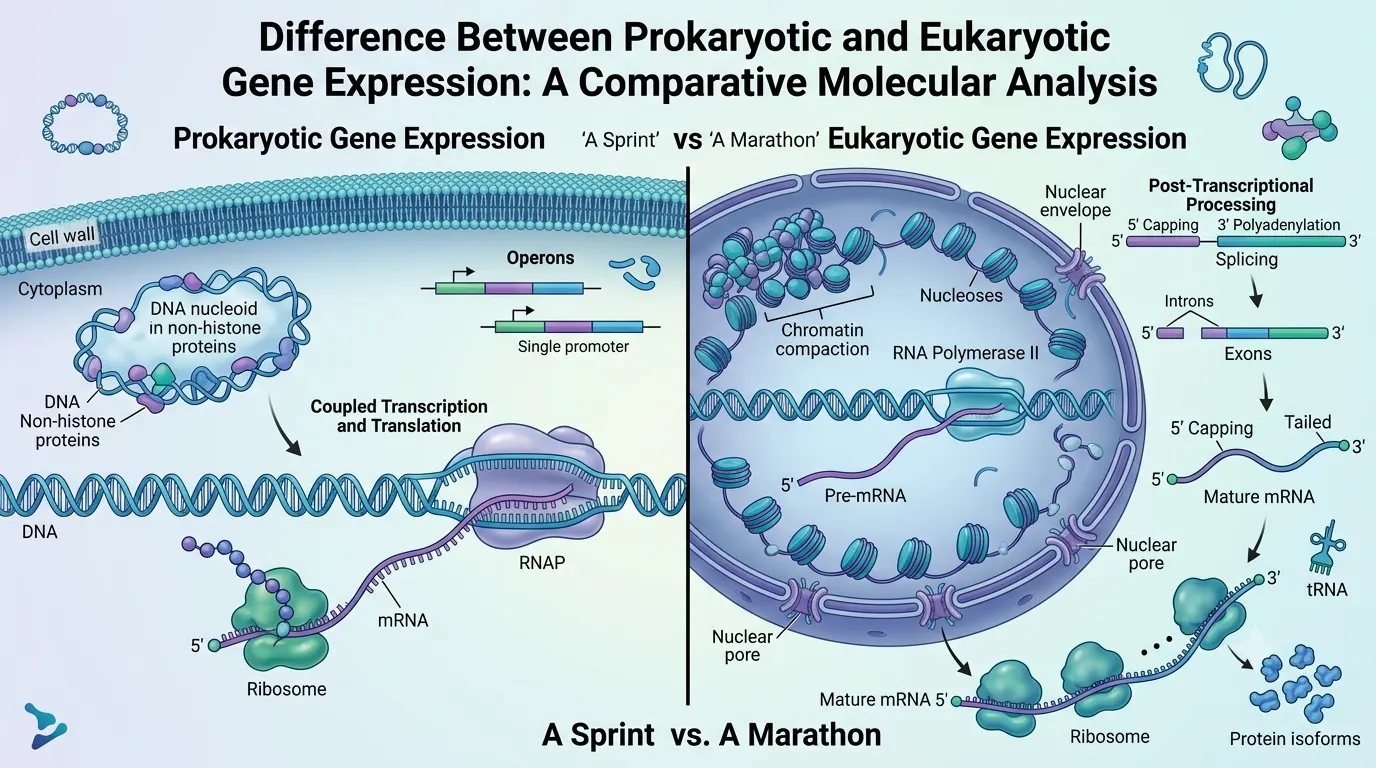
In the hierarchy of biological complexity, the transition from prokaryotic to eukaryotic life is most profoundly reflected in the architecture of gene expression. For the academic biologist, gene expression is not a simple linear flow from DNA to protein, but a multi-dimensional regulatory landscape. Understanding the divergence between these two systems is essential for applications in synthetic biology, where human genes are often "transplanted" into bacterial hosts, requiring a comprehensive "re-coding" of the genetic instructions.
Spatial and Temporal Compartmentalization
The defining divergence between prokaryotes (Bacteria and Archaea) and eukaryotes (Eukarya) is the presence of the nuclear envelope. In prokaryotes, the lack of a membrane-bound nucleus means that the genomic DNA is in direct contact with the cytoplasm. This leads to coupled transcription and translation. As RNA polymerase (RNAP) synthesizes an mRNA transcript, ribosomes immediately bind to the emerging 5' end and begin protein synthesis. This allows for near-instantaneous physiological responses to environmental stimuli.Eukaryotes, however, utilize spatial decoupling. Transcription and pre-mRNA processing occur within the protective confines of the nucleus, while translation is sequestered in the cytoplasm. This separation introduces a "checkpoint" system. mRNA must be fully processed and exported through the nuclear pore complex (NPC) before translation can begin. This delay allows for an additional layer of regulation—post-transcriptional control—which is virtually absent in prokaryotes.
Transcriptional Machinery and Initiation
The molecular tools used for transcription differ significantly in complexity. Prokaryotes utilize a single type of RNA polymerase, a multi-subunit enzyme that associates with a Sigma factor to recognize specific promoter sequences (like the -10 and -35 boxes). Regulation is often achieved through "operons"—groups of functionally related genes under the control of a single promoter, resulting in polycistronic mRNA.Eukaryotes employ at least three distinct RNA polymerases (Pol I for rRNA, Pol II for mRNA/snRNA, and Pol III for tRNA). Recognition of the eukaryotic promoter (often containing the TATA box) requires a massive assembly of General Transcription Factors (GTFs), such as TFIID and TFIIH, forming the Pre-Initiation Complex (PIC). Furthermore, eukaryotic gene expression is heavily influenced by "Distal Regulatory Elements" like enhancers and silencers, which can be located hundreds of thousands of base pairs away from the gene, communicating through DNA looping mediated by the Mediator complex.
Post-Transcriptional Processing: The Splicing Frontier
Perhaps the most technically complex aspect of eukaryotic gene expression is the "maturation" of the transcript. Prokaryotic mRNA is generally used in its "raw" form. Eukaryotic pre-mRNA, however, undergoes three critical modifications:5' Capping: The addition of a 7-methylguanylate cap, essential for ribosomal recognition and protection against exonucleases.
3' Polyadenylation: The addition of a Poly-A tail by Poly-A Polymerase, which dictates the half-life and stability of the transcript.
RNA Splicing: Eukaryotic genes are mosaic, composed of exons (coding) and introns (non-coding). The spliceosome, a complex of snRNPs, removes introns and ligates exons. Through alternative splicing, a single eukaryotic gene can produce multiple protein isoforms, vastly expanding the proteomic diversity of the organism. This explains how the human genome, with only ~20,000 genes, can produce over 100,000 different proteins.
Epigenetic Regulation and Chromatin Dynamics
Prokaryotic DNA is relatively "naked," although it is compacted by nucleoid-associated proteins. Eukaryotic DNA, conversely, is intricately wrapped around histone octamers to form nucleosomes. The degree of compaction (Euchromatin vs. Heterochromatin) is a master regulator of gene expression. Academic research at the doctoral level often focuses on the "Histone Code"—the specific patterns of acetylation, methylation, and phosphorylation that either "open" or "close" a gene for transcription. This epigenetic layer adds a fourth dimension to gene regulation that is far more sophisticated than the simple repressor/activator models found in bacteria.Conclusion
While prokaryotic gene expression is a sprint optimized for speed and metabolic efficiency, eukaryotic gene expression is a marathon of precision and complexity. For the biotechnologist, these differences mean that expressing a human gene in E. coli requires more than just the gene sequence; it requires the removal of introns (using cDNA), the addition of a bacterial promoter, and the optimization of codons to match the bacterial tRNA pool.References:
- Alberts, B., et al. (2017). Molecular Biology of the Cell (6th ed.). Garland Science.
- Kornberg, R. D. (2007). The molecular basis of eukaryotic transcription. Proceedings of the National Academy of Sciences, 104(32), 12955-12961.
- Lodish, H., et al. (2021). Molecular Cell Biology (9th ed.). W. H. Freeman.


No comments yet. You can leave the first one.