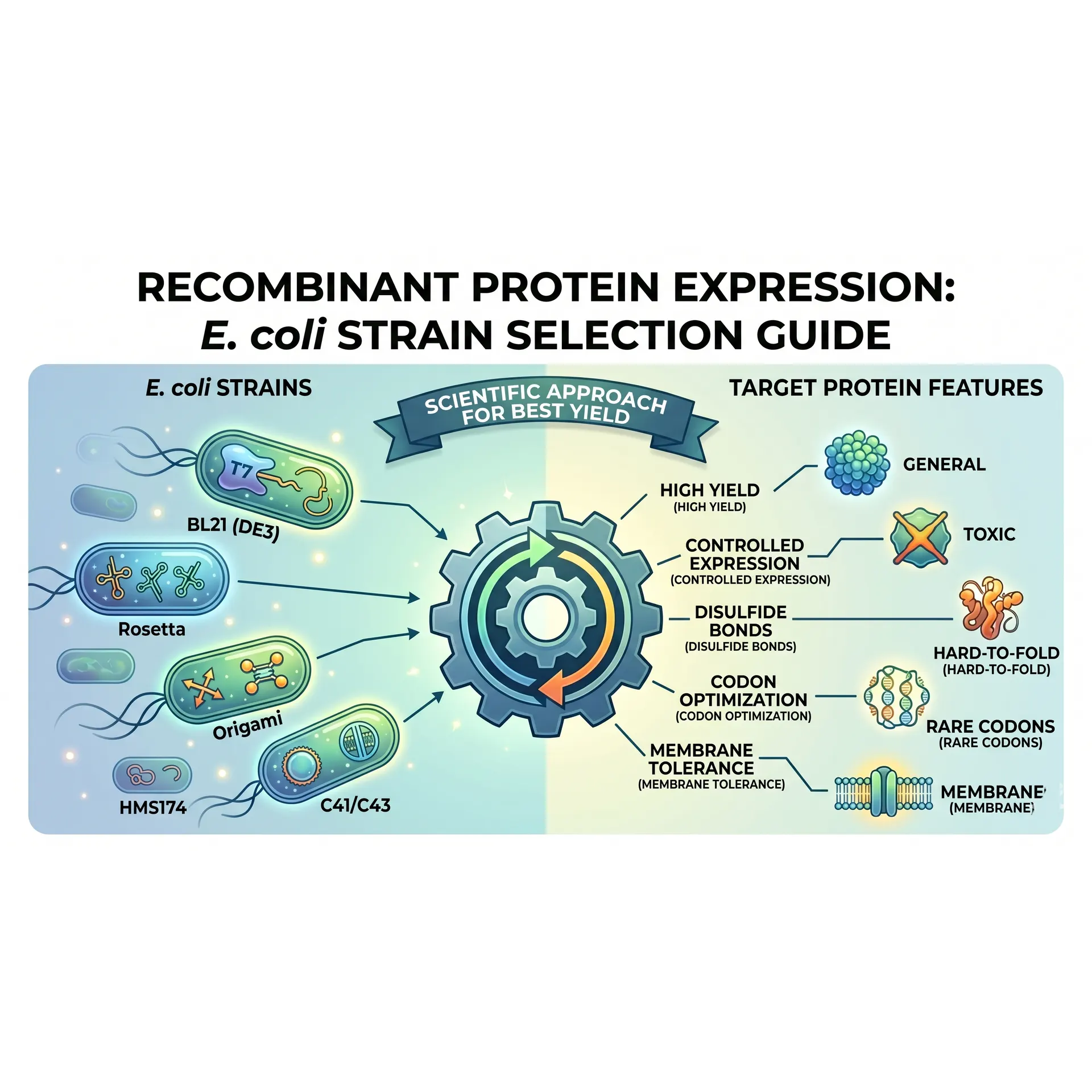
The unwritten first rule of surviving a master's program in a molecular biology lab is simple: not all Escherichia coli strains are created equal. The reliable DH5\alpha or Top10 cell lines you routinely use to transform your ligation products with delicate care will result in absolute disappointment if tasked with protein expression. Cloning strains are evolutionarily tailored to safeguard and replicate plasmid DNA; expression strains, conversely, are genetically engineered factories optimized exclusively for translation.
To harvest your recombinant protein with high yields and in a soluble, biologically active form, you must decode the genetic abbreviations listed in commercial strain catalogs. Choosing the wrong host can turn weeks of flawless cloning effort into insoluble, inactive inclusion bodies overnight.
Let's dissect the genetic markers behind the petri dishes on your laboratory bench and select the perfect E. coli worker for your target protein.
The Core Factory: BL21 and BL21(DE3)
If your target protein is a stable, "well-behaved" protein that requires no complex post-translational modifications or rare codon optimizations, your first line of defense is the BL21 lineage.
- Why BL21? Derived from the E. coli B strain, this line is genetically deficient in two major endogenous proteases: Lon (a cytoplasmic protease) and OmpT (an outer-membrane protease). This dual deficiency prevents your freshly synthesized recombinant protein from being degraded (proteolysis) during growth or downstream cell lysis.
- Decoding "DE3": If your expression vector relies on a T7 promoter system (such as the standard pET vector series), standard BL21 will yield zero protein. You require BL21(DE3). The "DE3" designation indicates that the host genome carries a stable lambda prophage lysogen encoding the T7 RNA Polymerase gene under the control of a lacUV5 promoter. When you introduce IPTG to the culture, the host synthesizes T7 RNA polymerase, which selectively targets the T7 promoter on your plasmid, launching transcription at extraordinary speeds.
Taming Toxic Proteins: BL21 pLysS and Tuner Strains
A common hurdle for graduate students is discovering that their target protein is toxic to E. coli. The culture grows normally during the lag phase, but upon IPTG induction—or even prior to it—the cells lyse and die. This is driven by "leaky expression," where the T7 promoter transcribes basal levels of the toxic gene before the culture reaches its optimum optical density (OD600).
- BL21 pLysS: This host carries a small, compatible plasmid (pLysS) that continuously expresses low levels of T7 lysozyme. This enzyme physically binds to and inactivates any basal, leaking T7 RNA polymerase molecules, completely silencing gene expression prior to induction. Once a massive dose of IPTG is introduced, the basal threshold is overwhelmed, allowing for tightly controlled, explosive protein production.
- Tuner(DE3): When you need to modulate expression levels gradually—akin to a valve rather than an "all-or-nothing" switch—Tuner is your ideal tool. It features a mutation in the lactose permease (lacY) gene, which regulates IPTG uptake. Instead of a subset of cells absorbing all available IPTG actively, Tuner allows uniform, linear diffusion of IPTG across the entire population. By adjusting the IPTG concentration micromolarly, you can slow down synthesis to prevent toxicity or misfolding.
Resolving Codon Bias: Rosetta and BL21 CodonPlus
Expressing eukaryotic genes (such as human, plant, or viral proteins) in a prokaryotic host often leads to a phenomenon known as Codon Bias.
The human genome regularly utilizes codons for arginine, leucine, isoleucine, proline, and glycine (e.g., AGG/AGA for Arg, AUA for Ile) that are extremely rare within the endogenous E. coli tRNA pool. When the bacterial ribosome encounters these rare codons, it stalls while waiting for scarce tRNAs, leading to translation arrest, truncated (incomplete) proteins, or amino acid misincorporation.
- Rosetta(DE3): This strain carries a specialized chloramphenicol-resistant plasmid supplying tRNAs for six rare codons (AUA, AGG, AGA, CUA, CCC, GGA). If your gene sequence was cloned directly from a eukaryotic cDNA library without prior codon optimization, Rosetta is an absolute necessity for rescuing your expression yields.
- BL21 CodonPlus: Similarly eliminates translation bottlenecks by providing extra copies of argU, ileY, and leuW tRNA genes, stabilizing translation elongation rates for AT- or GC-rich targets.
Disulfide Bonds and Soluble Folding: Origami and SHuffle Strains
High yields are meaningless if your target enzyme or antibody fragment accumulates as an inactive, misfolded aggregate inside an inclusion body. Many eukaryotic proteins require disulfide bonds (S-S) between cysteine residues to lock into their active, three-dimensional conformations. However, the wild-type E. coli cytoplasm is a highly reducing environment maintained by active metabolic pathways, meaning disulfide bonds cannot form stably.
Origami(DE3): This line features null mutations in both thioredoxin reductase (trxB) and glutathione reductase (gor) genes. Disrupting these pathways shifts the cytoplasmic redox potential to a more oxidative state, allowing disulfide bonds to form dynamically during translation.
SHuffle: Engineered by New England Biolabs, SHuffle represents a major upgrade to the Origami framework. In addition to trxB/gor mutations, it is constitutively modified to express a cytoplasmic version of DsbC (disulfide bond isomerase), an enzyme naturally restricted to the periplasm. SHuffle does not simply allow disulfide bonds to form; DsbC actively chaperones them, isomerizing mismatched links until the protein achieves its native, soluble confirmation. It is the premier choice for complex enzymes and single-chain antibody fragments (scFvs).
Evaluate the Promoter Architecture
Verify your plasmid configuration. If using a T7 promoter (pET series), you must select a host carrying the DE3 lysogen. For araBAD or tac promoters, standard BL21 or Tuner lineages are suitable.
Assess Codon Optimization Status
Analyze the gene sequence for rare prokaryotic codons. If expressing wild-type eukaryotic cDNA without codon optimization, route your choice to Rosetta or CodonPlus to avoid truncated products.
Map Structural Disulfide Bonds
Map Structural Disulfide Bonds:Conformation Check.Count the target cysteines. If your protein requires complex structural disulfide networks to remain functional and soluble, bypass standard lines and choose SHuffle or Origami.
Monitor Host Viability
If pilot expressions result in poor post-induction growth or premature cell death, implement pLysS to repress basal leaky expression, or drop temperatures to 18°C using Tuner to slow kinetics.
References
- Bhatwa, A., Jens, W., Dunnett, P., & van Dijl, J. M. (2021). Optimization of recombinant protein production in Escherichia coli. International Journal of Molecular Sciences, 22(14), 7432. https://doi.org/10.3390/ijms22147432
- Burgess-Brown, N. A., Sharma, S., Sobott, F., Loenarz, C., Oppermann, U., & Gileadi, O. (2008). Codon optimization can improve expression of human genes in Escherichia coli: A multi-gene study. Protein Expression and Purification, 59(1), 94-102. https://doi.org/10.1016/j.pep.2008.01.008
- Gopal, G. J., & Kumar, A. (2013). Strategies for the production of soluble recombinant proteins in Escherichia coli. International Journal of Cell Biology, 2013, 1-11. https://doi.org/10.1155/2013/919504
- Lobstein, J., Emrich, C. A., Jeans, C., Faulkner, M., Riggs, P., & Berkmen, M. (2012). SHuffle, a novel Escherichia coli protein expression strain capable of correctly folding disulfide-bonded proteins in its cytoplasm. Microbial Cell Factories, 11(1), 56. https://doi.org/10.1186/1475-2859-11-56
- Prinz, W. A., Aslund, F., Holmgren, A., & Beckwith, J. (1997). The role of the thioredoxin and glutaredoxin pathways in reducing disulfide bonds in the Escherichia coli cytoplasm. Journal of Biological Chemistry, 272(25), 15661-15667. https://doi.org/10.1074/jbc.272.25.15661
- Rosano, G. L., & Ceccarelli, E. A. (2014). Recombinant protein expression in Escherichia coli: Advances and challenges. Frontiers in Microbiology, 5, 172. https://doi.org/10.3389/fmicb.2014.00172
- Schumann, W., & Ferreira, L. C. (2004). Production of recombinant proteins in Escherichia coli. Genetics and Molecular Biology, 27(3), 442-453. https://doi.org/10.1590/S1415-47572004000300022
- Singha, T. K., Gulati, P., Antony, A., & Kapoor, V. (2017). Insights into T7 RNA polymerase compatible expression systems in Escherichia coli for recombinant protein production. Journal of Genetic Engineering and Biotechnology, 15(2), 293-299. https://doi.org/10.1016/j.jgeb.2017.07.009
- Studier, F. W. (2005). Protein production by auto-induction in high-density shaking cultures. Protein Expression and Purification, 41(1), 207-234. https://doi.org/10.1016/j.pep.2005.01.016
- Terpe, K. (2006). Overview of bacterial expression systems for heterologous protein production: From molecular biology to commercialized product. Applied Microbiology and Biotechnology, 72(2), 211-222. https://doi.org/10.1007/s00253-006-0465-8
No comments yet. You can leave the first one.